
Yfull Interpretation service

One of the many good companies
HAPLOGROEPEN
Y en Mt

Whole Genome Sequencing - 100% of DNA
YSEC DNA shop
|
Yfull Interpretation service |
One of the many good companies |
HAPLOGROEPEN
|
Whole Genome Sequencing - 100% of DNA |
YSEC DNA shop |
|
* |
|
* De Engelstalige pagina's zijn vollediger. |
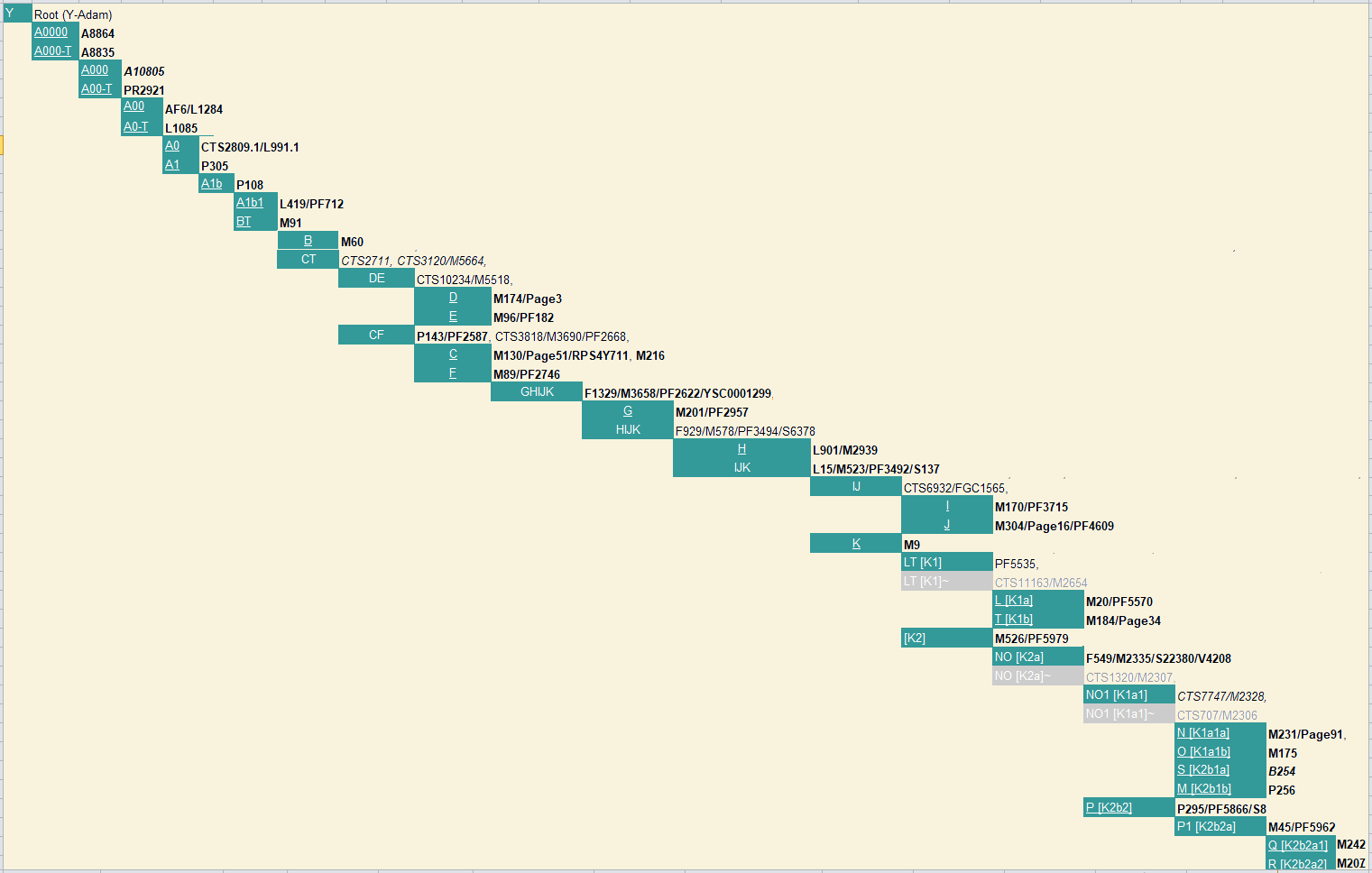
Y-Tree-ISOGG van Ray Banks. (17) |
|
* 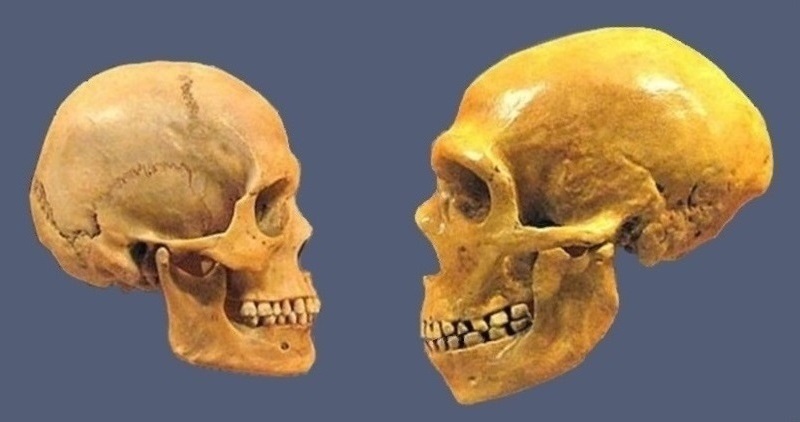
Schedel van een moderne mens en van een Neanderthaler, Museum of Natural History in Cleveland. (15) De mensheid wordt stamt uit de Homo Heidelbergensis, ook bekend als Homo Rhodesiensis, een soort of ondersoort van archaïsche mensen in het geslacht Homo dat in het Midden Pleistocene - circa 700.000 is uitgestorven. Denisovans en Neanderthalers splitsten zich ongeveer 750.000 jaar geleden van Homo sapiens af en gingen ongeveer 100.000 jaar later uit elkaar. * 
Reconstructie van Homo erectus uit Georgië (16a) * DenisovansA0000 - A8864 → A8983 * Neanderthalers |
|
A000T
|
A8835
|
|
* Homo Sapiens SapiensDe oudste gemeenschappelijke menselijke voorvader is een man aan wie de code A00 is gegeven. Directe afstammelingen van hem leven in Kameroen in Afrika. Men acht de tijd tot de meest recente gemeenschappelijke voorvader van de gehele mensheid (TMRCA) 250.000 jaar (plus of min 50.000 jaar). (16) De oudste archaeologisch gevonden menselijke fossielen zijn gevonden in Jebel Irhoud in Marokko. Ze worden 315 ±34 duizend jaar oud gedateerd. Alle mensen in de wereld stammen af van een vrouw waarvan we denken dat zij rond de 200.000 jaar geleden in Afrika leefde. Dat dit veel jonger is komt doordat de nakomelingen van eerdere vrouwen uitgestorven zijn. Alle Y-DNA haplogroepen zijn ontsproten uit de gemeenschappelijke stamvader van alle mensen. Hij leefde in Afrika en hij had haplogroep A00-AF4. * Y-DNA Haplogroep A00-AF6/L1284.
Y-DNA Haplogroup BT-M91, Hieruit komen de Haplogroepen B-M60 en haplogroep CT-CTS2711. Zij zijn de voorouders van alle Euraziatische volkeren. . Y-DNA Haplogroep B-M60 ontstond in Afrika ongeveer 50.000 jaar geleden. Hij is van onbekende herkomst. Deze man en zijn nakomelingen bleven in Afrika. Deze leven daar nu nog in geringe concentraties verspreid onder de jager-verzamelaars van Ethiopië en Soedan en bij de kliktaal sprekers.
Y-Y-DNA Haplogroep CT-M168, CTS2711, is afkomstig van BT-M91. CT-M168* is nooit gevonden, maar afstammelingen van Haplogroup CT treffen we aan in verschillende prehistorische menselijke fossielen die op oud DNA onderzocht zijn, inclusief specima die zijn geassocieerd met het Pre-Pottery Neolithic C (1/1; 100%), Neolithisch Ganj Dareh Iran (1/2; 50%), Natufian (2/5; 40%), Pre-Pottery Neolithic B (2/7; ~ 29%), Alföld Linear Pottery (1/1 op twee ALP archeologische vindplaatsen; 100% ), Lineair bandkeramiek culturen (1/2 op Karsdorf LBK archeologische site, 50%) en enkele Upper Paleolithische Europeananen (Cioclovina1, Kostenki12, Vestonice13). Maar of deze, dan wel of ze allemaal behoren tot paragroep CT * of tot zijn subgroepen, is nog niet bepaald.
Y-DNA Haplogroep CF-P143 is gesproten uit CT-M168, het de broeder-haplogroep van DE-CTS10234. Uit CF komen de haplogroepen C-M130 end F-M89. Het bestaan en de verspreiding van deze clade wordt afgeleid uit het feit dat haplogroepen die afstammen van CF de meeste mannelijke afstammelingslijnen in Eurazië, Oceanië en Noord- en Zuid-Amerika omvatten. Y-DNA Haplogroep C-M130 komt voort uit CF-P143, that stamt van CT-CTS2711, en is kort nadat de moderne mens in Azië aangekomen was ontstaan, ongeveer 90.000 jaar geleden. Nakomelingen volgden een route over het Zuid-Arabische schiereiland, door Pakistan en India tot Sri Lanka, naar Zuidoost Azië en Japan, naar Polynesië en Australië.
Y-DNA Haplogroep DE-CTS10234, stamt tezamen met zijn broedergroep CF-P143 uit CT-CTS2711, de leeftijd van haplogroep DE wordt over het algemeen geschat tussen 65.000 en 71.000 jaar. DE is nu in verschillende clusters over Eurazië verspreid. Van DE komen de haplogroepen D-M174 en E-M96. Y-DNA Haplogroep D-M174 ontstond omstreeks 50.000 jaar geleden en komt hoofdzakelijk voor in Centraal en Zuidoost Azië en op Japan.
|
|
Y-DNA Haplogroep E-M96 komt voort uit haplogroep B. Ontstond in Noordoost Afrika ongeveer 40,000 jaar geleden, en verspreidde zich over dit hele continent met de Bantoe landbouwcultuur. E is ook de meest voorkomende afstamming onder Afro-Amerikanen. Het is een oude, diverse haplogroep met vele takken en is nu over geheel Afrika verspreid. Het is in kleine aantallen in Noord-Afrika en het Midden-Oosten Het behoorde bij de eerste emigraties van moderne mensen naar buiten Afrika. Zijn subgroep E1b1 heeft later in Afrika, maar ook daarbuiten de verste vertakkingen en de grootste verspreiding gekregen. |
|
|
E1a1-M44, is in meer dan de helft van de Fulbe bevolking in Kameroen, maar slechts in 2-5% in Mali en Soedan, ontstond 19.000 jaar geleden. In Elzas-Lotharingen.
Y-DNA Haplogroep F-M89 stamt stems from CF-P143 en is de stamvader van alle Y-DNA haplogroepen van G tot T. Zijn nakomelingen omvatten nu 90% van de wereldbevolking. Hij kan 60.000 tot 80.000 jaar geleden geleefd hebben.
Y-DNA Haplogroup GHIJK-F1329, stems from F-M89, zijn subgroepen omvatten de overgrote meerderheid van de mannelijke bevolking van de wereld met de haplogroepen G, I, J, K, L, M, N, O, P, Q, R, S and T. Y-DNA Haplogroep G-M201 stamt via F2-M427 en F1329 uit F-M89, en is omstreeks 45.000 jaar geleden ontstaan in het Midden-Oosten. Haplogroep G komt in Europa bij 3 tot 7% van de mannen voor. Komt wat meer voor in de Alpen, en oplopend tot 10% op de eilanden Sardinië en Corsica. Deze haplogroep is in het begin van het Neolithicum Europa binnengekomen. Sommige subgroepen kwamen mogelijk later en tezamen met J1 en/of J2. G wordt relatief vaak gevonden in archeologisch DNA, daarna veel minder.
* De eerste Europese landbouwers hadden G2a De mannen van de eerste menselijke nederzettingen in Europa hadden G2a-P15 en G2a2b-L30. Het is gevonden in een neolithische begraafplaats bij Derenburg in Saxen-Anhalt (Duitsland). Het wordt gedateerd tussen 7.000 en 7.500 jaar geleden.
Rond 7000 jaar geleden arriveren deze eersten in onze contreien. Ze vestigden zich op de berghellingen van het Zuid-Limburgse Maasdal, zoals op Caberg een wijk aan de rand van het plateau ten westen van Maastricht. Hier bevindt zich de vruchtbare lössgrond. Bouwstenen vinden ze in de ondergrondse mergel met vuursteen voor de fabrikage van werktuigen. Hun aardewerkpotten hebben de Bandkeramische stijl. De cultuur bloeit gedurende 500 jaar om dan spoorloos te verdwijnen. In hoeverre met deze cultuur de residente bevolking verdween dan wel bij de nieuwkomers is opgenomen verschillen de opvattingen. 
Impressie van een Bandkeramisch dorp te Caberg bij Maastricht. (19) * Y-DNA Haplogroep HIJK-F929, hieruit komen de haplogroepen H-M69, IJ-FGC1665 and K-M9. Y-DNA Haplogroep H-M69 Ontstond bij een F-man zeer waarschijnlijk in India 30.000 tot 40.000 jaar geleden. Bijna alle dragers leven in India. In Europa komt het voor bij Zigeuners, ook wel Roma of Sinti genoemd, die naar aangenomen wordt uit India komen.
Y-DNA Haplogroep IJK-L15 ontstond uit HIJK - F929/M578, hieruit stammen IJ-FGC1665 en K-M9. Y-DNA Haplogroup IJ-FGC1665 , hieuit ontstonden de oudste volkeren van Europa (vooral in Scandinavië en de Balkan), Anatolië, het Midden Oosten (met name Arabië, de Kaukasus, Levant en Mesopotamië) en de kusten van Noord-Afrika. Als gevolg van de massale migraties in latere tijden zijn ze nu ook te vinden in Noord- en Zuid-Amerika en Australë. Y-DNA Haplogroep I-M170 ontstaan 43.000 jaar geleden is een Europese groep. Hij wordt daar gevonden in archeologischeresten. I* is gevonden in skeletten uit het Gravettien uit Krems-Wachberg, Oostenrijk, gedateerd op 31.000 cal BP.. In een recente studie werd van 5 mannen het Y-DNA bepaald uit het mesolyticum allen hadden I, I-P38+ (2x),en I2a1b-CTS176 (3x). Dit suggereert een ontstaan in Europa waar hij waarschijnlijk voor het laatste glaciale maximum is ontstaan. Hij komt hier bij ongeveer een vijfde deel van de bevolking voor. Buiten Europa nauwelijks.
I2-M438 zien we in Armenië, Georgië en Turkije.
|
Y-DNA Haplogroep I1a |
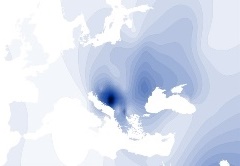
Y-DNA Haplogroep I1b |
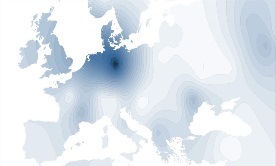
Y-DNA Haplogroep I1c |
|
Haplogroep J-P209 ontstond in Voor-Azië en ging vandaar naar Noord Afrika Europa, Centraal Azië, Pakistan en India. Het meeste komt J voor in het Midden-Oosten. De aanwezigheid daalt geleidelijk in de richting naar Noordwest Europa, waar de kustbewoners 3% hebben. De verspreiding vond zowel in het Neolithicum plaats als ook door periodieke latere immigratiegolven. J1-L255/M267/PF4646 subgroepen kunnen een meer zuidelijke oorsprong hebben omdat zij vaker in de Levant en andere delen van klein Azië en Noord Afrika gezien worden. Er is een geringe verspreiding in de zuidelijke Méditerranée en in Ethiopië. J2-L228/M172/S321 lijnen ontsprongen in de regio van de Vruchtbare Halve Maan. De belangrijkste verspreiding vond plaats naar de Mediterrane regio, waarschijnlijk in het Neolithicum. Het is onbekend wanneer J2 Centraal Azië, Pakistan en India bereikte.
|
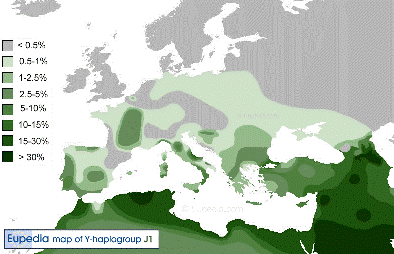
Y-DNA Haplogroep J1 |
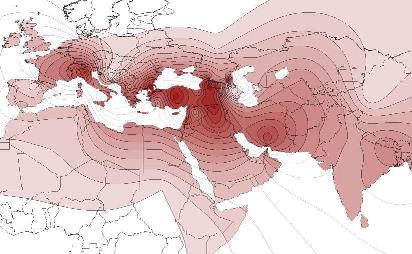
Y-DNA Haplogroep J2 |
|
< Y-DNA Haplogroup K-M9 is een oude lijn die ongeveer 50.000 jaar geleden ontstond, waarschijnlijk in Zuid of West-Azië. Er zijn twee takken: de eerste tak is opgesplitst in K1 tot en met K4, die in lage frequenties worden gevonden in Afrika, Eurazië, Australië en de eilanden van de Stille Oceaan. Uit de tweede tak ontsproten de haplogroepen L tot T.
Y-DNA Haplogroup LT-L298/P326, PF5535 (former K1) splitst zich in: L-M20 en T-M184. Y-DNA Haplogroep L-M20 verdeelt zich in:
Y-DNA Haplogroep M-P256 komt alleen in Papoea Nieuw Guinea voor en daar bij twee derde van de bevolking. Y-DNA Haplogroup N-M231 komt verspreid voor van Zuidoost-Azië tot Oost-Europa. De vroegste vondst in Europa is uit de IJzertijd in Hongarije, maar is hij wordt hier nu niet meer aangetroffen.
Y-DNA Haplogroep O-M175 is waarschijnlijk in Zuid-China ontstaan en verspreidde zich van daar over China, Taiwan, Indonesië en de eilanden van de Stille Oceaan.
Y-DNA Haplogroup P-P295, vroegerK2b2, is ongeveer 35.000 jaar geleden ontstaan uit K in Centraal-Azië en is nu te vinden in Oezbekistan, Kazachstan en Zuid-Siberië. Hieruit ontstonden de haplogroepen Q en R, de dominante haplogroepen van Europa en nu ook van Amerika.
Y-DNA Haplogroep Q-M242 ontstond 20.000 jaar geleden in Centraal Azië en verspreidde zich via de Altai/Baikal regio in Noord Eurazië naar China en ging omstreeks 16.000 via de Beringstraat naar Amerika. Het wordt in zijn ontstaansregio nog gevonden en bij enkelen in Europa.
Y-DNA Haplogroep R-M207/Y482 ontstond rond 28.000 jaar geleden in Centraal Azië uit F.
R1-M173/Y465. deze heeft twee grote subgroepen: R1aL146/M420 en R1b-M343/PF6090. Y-DNA Haplogroep R1a-L146/CTS2443, ontstaan 22.800 jaar voor heden, TMRCA 18.200 jaar voor heden ook deze tak ontsprong zeer waarschijnlijk op de Eurazische steppe en komt nu het meest voor in Oost-Europa en in West- en Centraal-Azië.
R1b-M343/PF6242, ontstaan 22800 jaar voor heden, TMRCA 20400 jaar voor heden. De oudste vormen van R1b worden in zeer lage frequenties van West-Europa tot in India gevonden in een uitgestrekt gebied waar de nomadische R1b-jagers-verzamelaars tijdens de ijstijd rondzwierven. De twee oudste archeologische vindplaatsen die tekenen van vee-domesticatie vertonen zijn de dorpen Çayönü Tepesi in het zuidoosten van Turkije en Dja'de el-Mughara in het noorden van Irak, op 250 km afstand van elkaar. Dit is vermoedelijk het gebied van waaruit zij zich begonnen uit verspreiden en is naar we aannemen het oorspronkelijke thuisland van R1b.
R1b1a2-M269/S13 bleef aanvankelijk in de Noord-Kaukasus en de Pontische Steppe tussen de Dnjepr en de Wolga; wordt gevonden in Europa, voornamelijk West-Europa, maar ook in Zuidwest-Azië. Deze clade is nauw verbonden met de verspreiding van de Indo-Europese talen, zoals blijkt uit de aanwezigheid in alle delen van de wereld waar in de oudheid Indo-Europese talen werden gesproken; In Nederland heeft meer dan de helft van de bevolking deze subgroep. Hij ontstond 4000 tot 8000 jaar geleden in Zuidwest-Azië en verspreidde zich later naar Europa. De explosieve groei ging van (schijnbaar) nul naar een solide meerderheid in een groot deel van West-Europa gedurende de laatste paar duizend jaar; dit is waarschijnlijk een van de meest interessante gebeurtenissen in de recente Europese geschiedenis.
R2-M497/Y3316, ontstond 28.200 jaar geleden, TMRCA 16.300 jaar geleden, is in Centraal Azië en India. |
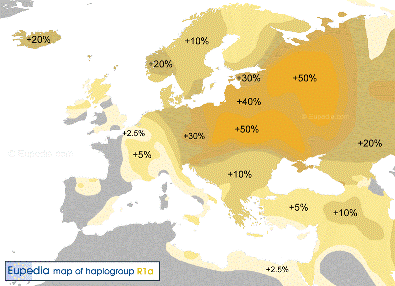
Y-DNA Haplogroep R1a |
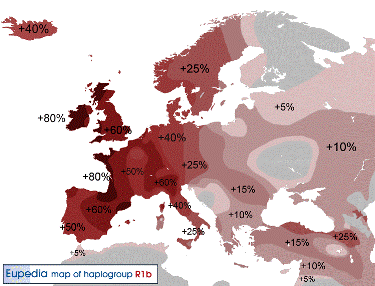
Y-DNA Haplogroep R1b |
|
Y-DNA Haplogroep S is de haplogroep van de hooglanden van Papoea Nieuw Guinea. Het komt daar bij de helft van de bevolking voor. In lage aantallen ook op naburige eilanden. Y-DNA haplogroep T-M184 Is een onmiddellijke afstammeling van de haplogroup LT, wiens ouderlijke clade haplogroup K is. De clade is 40.000 oud. Hij wordt gevonden in zijn hoogste frequenties bij sommige bevolkingen in Oost-Afrika en Oost-India, naar verwachting te wijten aan relatief recente migraties. Bij frequenties van meer dan 30% zien we hem bij Somaliërs van Djibouti, in Madagascar, Bauri en Yerukula van Oost-India, Argyns uit Kazachstan en in Sicilië.
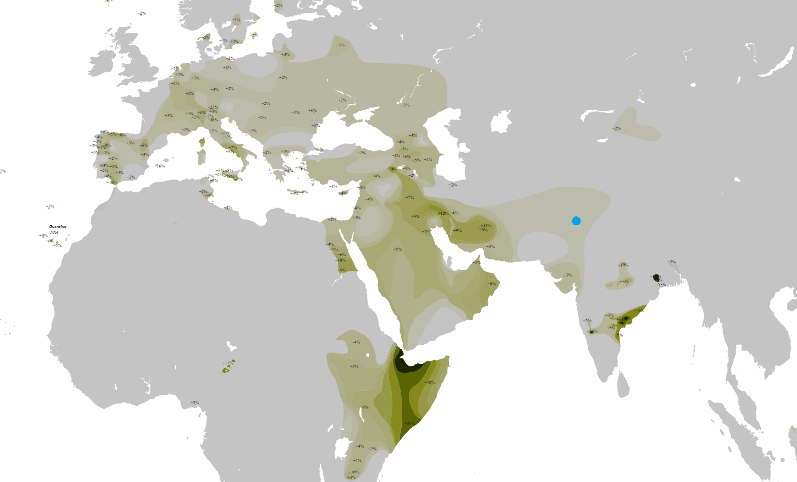
Y-DNA Haplogroep T * |
Twee stambomen van de mensheid |
|
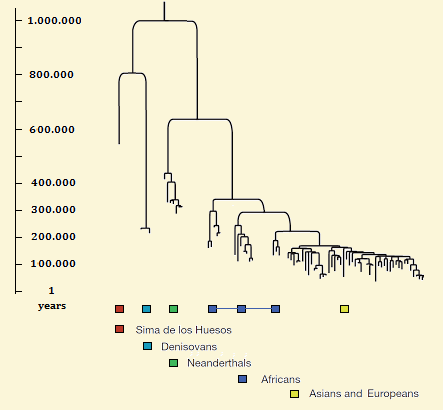
Homines boom (24) |
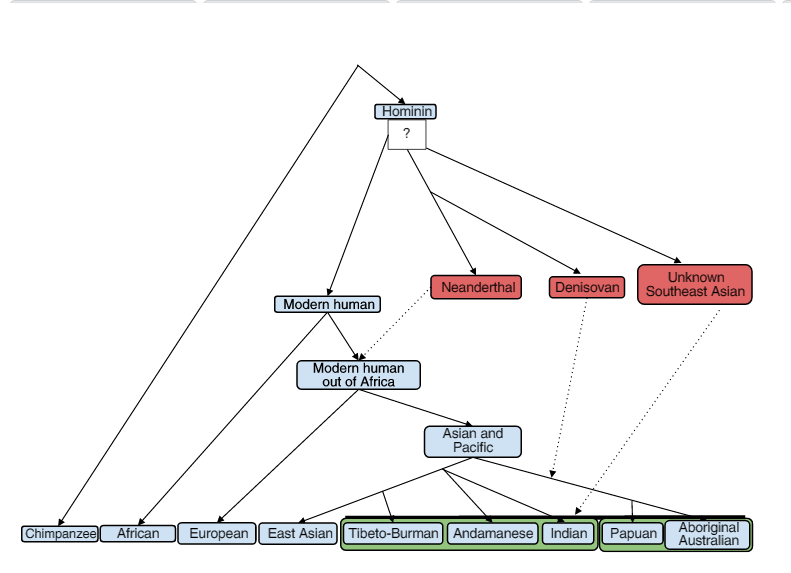
Stamboom van de hominiden. (24a) |
|
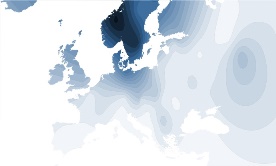
* Hypothetische Y-DNA verspreiding in Europa in de Pre-Romeinse Tijd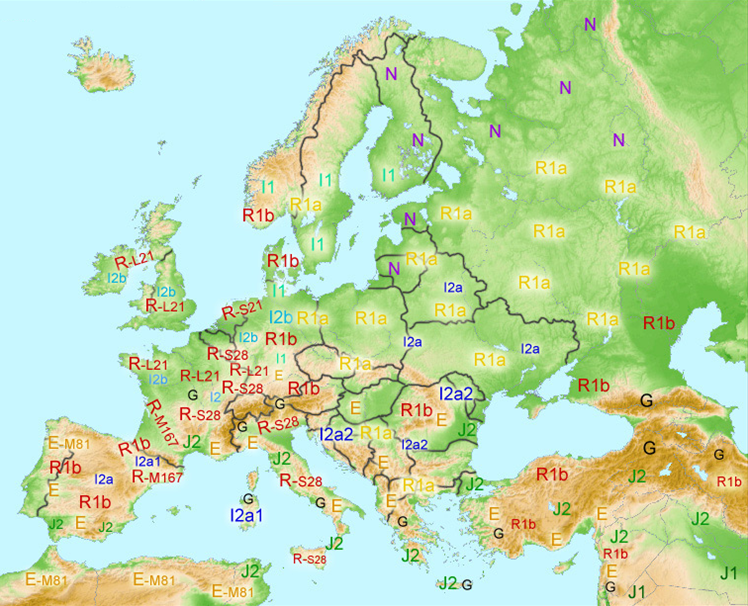
|
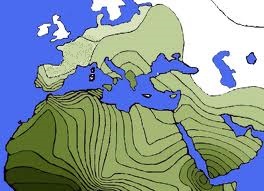
|
* Wereldkaart van Y-DNA Haplogroepen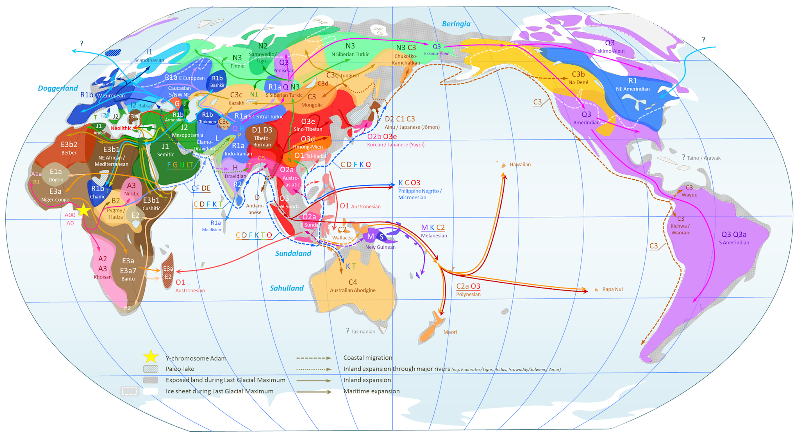
Beweeg de muis over de kaart voor een vergroting van West-Europa. (25) Deze kaart toont duidelijk aan hoe inheemse bevolkingen in de korte tijd van een paar eeuwen van de kaart worden geveegd. Een voorbeeld is de verspreiding van haplogroep G die vroeger domineerde in Europa maar daar nu niet meer getekend staat en enkel een haard heeft in de Kaukasus. In Noord Amerika is haplogroep Q verdrongen door het Europese R1. |
|
* mt-DNA HaplogroepenMitochondriaal DNA (mt-DNA) is een klein deel van het genoom - slechts ongeveer 1 / 200.000ste deel ervan - dat langs de moederlijn wordt doorgegeven van moeder naar kind en langs de dochters naar de kleinkinderen. De verspreiding van de mitochondriale haplogroepen vanuit Afrika (Jean Manco) 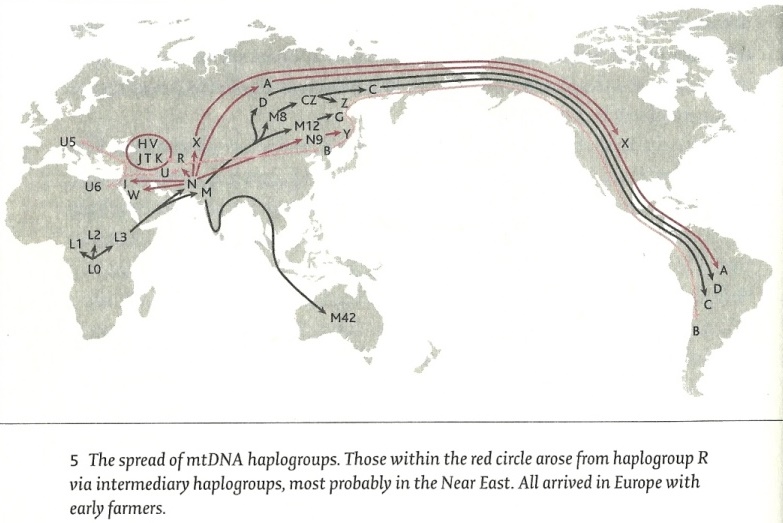
Door de sequentie van de volledige Y-chromosomen en de mitochondriale genomen van 69 mannen uit negen populaties is de tijd bepaald waarop de gemeenschappelijke voorouder van het mitochondriale genoom leefde. Deze MT-TMRCA wordt nu gesteld op 100.000 tot 150.000 jaar. (26) Een groot voordeel van mt-DNA is dat het in vergelijking tot Y-DNA aanzienlijk beter bewaard blijft in archeologische resten. Vaak kan dan op andere wijze zoals door de radiocarbon methode de ouderdom goed vastgesteld worden. Mt-Haplogroep L0 noemen we de configuratie van een vrouw die ongeveer 125.000 jaar geleden in Zuid- of Oost-Afrika leefde. Alle mitochondriale DNA lijnen van de huidige mensheid stammen van haar af
De vrouwen in de groepen die zestig tot tachtig duizend jaar geleden vanuit Afrika Eurazië binnenkwamen hadden de haplogroepen M en N. mt-Haplogroep M ontstond ongeveer 60 tot 80.000 jaar geleden, waarschijnlijk al in Noordoost Afrika maar mogelijk ook in Arabië en verspreidde zich over Azië en Europa. In Europa verdween deze groep tijdens het laatste glaciale maximum. Hierna bleek het vervangen door subcladen van N. Uit M ontstonden in Eurazië de haplogroepen C, Z, D, E, G en Q. Mt-Haplogroep N, ontstond evenals M rond 70.000 jaar geleden. Het is de mt-DNA haplogroep waaruit alle huidige Europeanen stammen. Deze groep is nog in Afrika ontstaan en komt nog steeds in uiterst lage frequenties verspreid over Europa en Azië voor. Zijn subgroepen ontstonden gedurende de laatste ijstijd toen wij nog jager-verzamelaars waren.
Uit N ontstonden de subgroepen O, A, S, R, I, W, X, Y, B, F, J, P, U, H, V, J, T en K. * Mt-Haplogroep C is waarschijnlijk tijdens het Mesolithicum verspreid vanuit een IJstijd refuge bij het Baikalmeer, en het dal van de Jenisej rivier, en de Altai en Sayan bergen. Deze haplogroep bestond onder pottenbakkers van het Baikalmeer ongeveer 7500 jaar geleden.
mt-Haplogroep pre-HV is geëxtraheerd uit een Cro-Magnon die 28.000 jaar geleden in Zuid-Italië stierf. Het is de voorloper van de groepen H en V. mt-Haplogroep H ontstond voor 35.000 jaar geleden in het Klein Azië. Hij is in Europa met 40% aanwezigheid de grootste groep. Opvallend is dat in archeologisch Europees DNA van 7.500 jaar geleden slechts de helft van dit percentage gevonden wordt en het in Mesolitisch materiaal nauwelijks aanwezig is.
(27)
Mt-haplogroep I ontstond ongeveer 20.000 jaar geleden in het Nabije Oosten of in de Kaukasus en verspreidde zich in golven. Een golf ging naar Noord-Europa, voornamelijk Noorwegen en Finland; een andere naar Pakistan en Noordwest India. Het is nog steeds aanwezig in de Oekraïne rond de Kaspische Zee, en in Anatolië en Griekenland. Het lijkt samen te hangen met het Proto-Indo-Europese cultuur, er lijkt verband te zijn met Y-DNA-R, vooral R1a. Sommigen denken dat het een van de oorspronkelijke Mt-haplogroepen van Europa is en later verdreven is naar het noorden en andere afgelegen plaatsen. Er zijn nog geen archeologische vondsten gedaan, maar wel in grotere hoeveelheden in meer recente historische begraafplaatsen in Scandinavië. Zijn latere verspreiding kan plaats gevonden hebben door de Vikingen. De Kroatische eiland Krk is een hotspot. mt-Haplogroep J is weer een oude haplogroep die 45.000 jaar geleden in Klein Azië ontstond. We nemen aan dat hij met de neolithische landbouwcultuur naar Europa kwam. Hij is algemeen in Centraal Azië en boven de Kaukasus. Hij is verbonden met de Indo-Europese cultuur en gaat samen met de Y-DNA haplogroep R1b.
mt-Haplogroep K stamt uit U8. Hij wordt door heel Europa en in West Azië tot in India gevonden. De hoogste concentratie ligt echter in Noordwest en Centraal Europa, in Anatolië en Zuid Saoedi-Arabië. Men denkt dat hij 16.000 jaar geleden in Egypte of Anatolië is ontstaan. Hij heeft ondanks zijn geringe ouderdom de meeste subgroepen.
K1a is de grootste subgroep. Het feit dat hij in Klein Azië zo veel voorkomt doet vermoeden dat hij er al voor de Neolithische expansie naar Europa was, temeer omdat hij in Europa voor de Neolithische intocht ontbrak en daarna plotseling in een percentage van 17% voorkomt, overigens ligt dat bij de huidige Europeanen weer lager, maar het is gelijk aan dat in de huidige Levant.
Mt-Haplogroep R ontstond uit N en hieruit kwamen deze 6 Europese mt-Haplogroepen pre-HV, T, U en K en de oosterse haplogroepen R1 tot R31 in Azië, Australië en in de Amerika's. mt-Haplogroep T is rond 15.000 jaar geleden in Mesopotamië of Noordoost Afrika ontstaan. We zien het in Noord Europa, Noord Afrika, Centraal Azië en Siberië met haarden in India en Noordwest China.
Mt-Haplogroep U ontstond 60.000 jaar geleden uit R. Dit is het enige archeologische Europese DNA dat tot 5.600 jaar geleden gevonden is. Rond 5000 jaar terug is het abrupt vervangen door de zeer grote diversiteit van de neolithische boeren.
(28)
mt-Haplogroep V ontstond waarschijnlijk op het Iberische schiereiland 15.000 jaar geleden onder de jager-verzamelaars die zich daar tezamen met h2, H3 en U5 mannen tijdens het laatste glaciaal hadden teruggetrokken. Bij de klimaatverbetering hebben ze Europa van daar uit weer herbevolkt. Nu is het weer een grote groep in Europa met zijn hoogste concentratie in Noord Scandinavië bij de Lappen 40% en van daaruit dalend in West Europa. In Nederland 8%. mt-Haplogroep W is 18.000 jaar geleden ontstaan. Zijn verspreiding is ongeveer gelijk aan die van mt-Haplogroep I. Zijn hoogste aanwezigheid is in de Oekraïne, Europees Rusland, de Baltische staten en Finland, maar overal ongeveer 3 tot 5%. In Noord-Pakistan 15%, Noord India 10%. Daar vooral in de hogere kasten en bij de Indo-Europese sprekers. Ook hier weer vaak samen met Y-DNA R1a en R1b. mt-Haplogroep X is ouder dan 30.000 jaar. In lage concentraties wordt het overal en in iedere bevolkingsgroep gezien. Hij komt voor in Europa, Noord Afrika en in Amerika bij de Indianen. De aanwezigheid is meestal 1 tot 2 % en komt zelden boven de 5%.
|
*
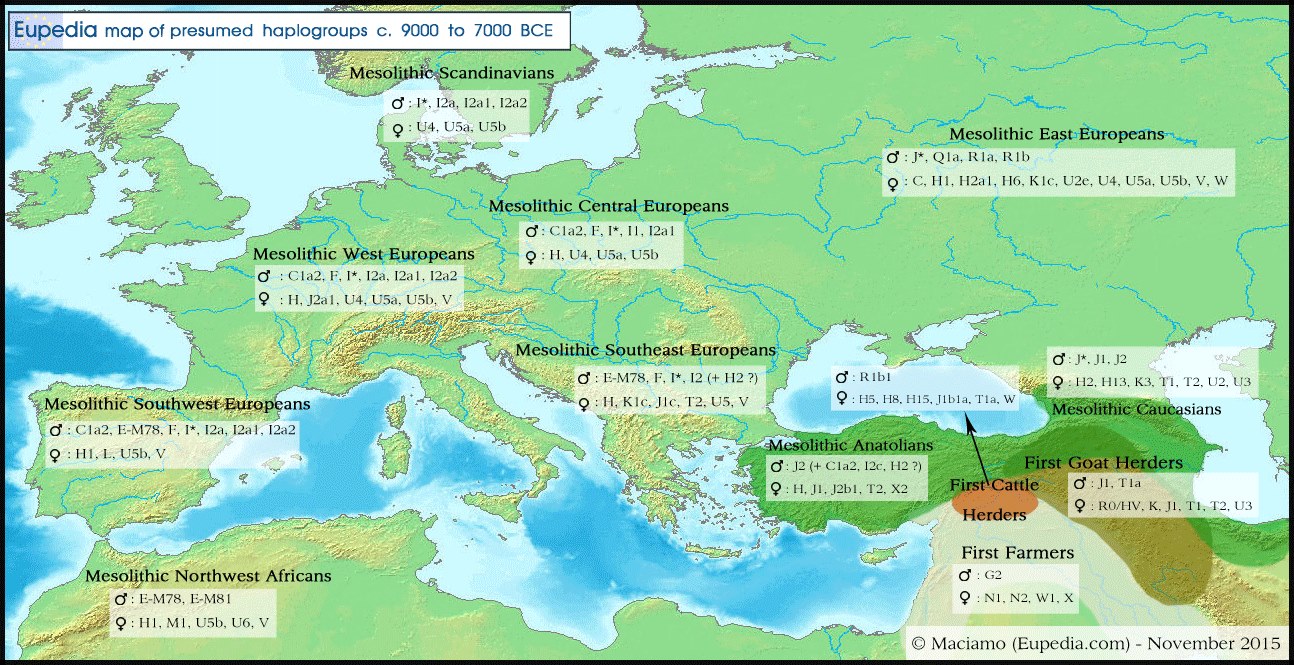
Verdeling van de Y-DNA en Mt-DNA haplogroepen in Eurazië van circa 9000 tot 7000 jaar geleden.
|
* Wat zijn wij in de NederlandenOnderstaande cirkeldiagrammen tonen de verdeling van de tien grootste Y-DNA haplogroepen die in Nederland bij 410 mannen in het jaar 2008, en in Vlaanderen in het Oud Hertogdom Brabant Project bij 1057 man in het jaar 2013 man zijn waargenomen. Dit is niet echt het verschil tussen België en Nederland maar het verschil tussen Nederland en het Nederlands sprekende deel van België: dus Vlaanderen, Antwerpen en beider provincies Brabant en Limburg. Er zijn verhoudingsgewijs weinig Walen in de Belgische groep. (32) |
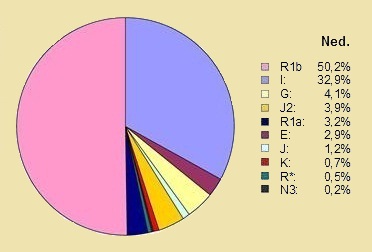
Nederland |
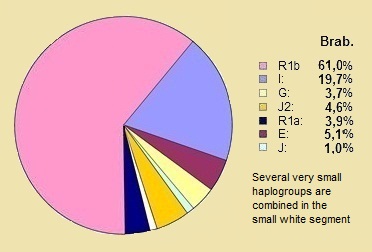
België |
Mesolitisch DNAHet oudste archeologisch gevonden Y-DNA is C-M130*. Het is gevonden in een 37,000 jaar oud skelet in Kostenki aan de oevers van de Don in Rusland. Van een Europese jager-verzamelaar die ongeveer 7000 jaar geleden in La Braña, Spanje, stierf kon ook mt-DNA, autosomaal DNA en Y-DNA bepaald worden. Uit het au-DNA bleek dat hij een donkere huidskleur en blauwe ogen had. Zijn Y-DNA haplogroep is C1a2-V20. Deze clade is nu in Europa zeer zeldzaam. |

|
|
Dit is een aanwijzing dat Europa ten tijde van de jager-verzamelaars bewoond werd door bruine mensen. De witte (roze) huidskleur kan door de neolithische boeren naar Europa zijn meegekomen. De landbouwbevolking had deze verkregen in Zuidwest Azië door genetische contacten met Neanderthalers. Deze waren meer dan tweehonderdduizend jaar eerder in Eurazië waren aangekomen en hadden hun pigmentatie daar zeer waarschijnlijk door natuurlijke selectie verloren in een continent met veel minder zonnestaling dan het land van herkomst, Afrika. De depigmentatie van de moderne mens heeft in Europa volledig plaats gevonden en in Zuid Azië, zoals India, veel minder of soms zelf niet. (33) Neolithisch DNAEen abrupte omslag is er 7200 jaar geleden bij de binnenkomst van de eerste neolithische boeren golf. Dit is gevonden in Duitsland en Hongarije, Bij hun domineert haplogroep G2a en zijn subgroepen G2a2b en G2a2b. De oudste Europese mummie, die gevonden werd in een gletscher in Tirol in het Ötzenthal en 5300 jaar geleden leefde had G2a2a1b1a1-FGC5672 en zijn mt-DNA haplogroep is K1. Een familie uit Thüringen in Unterfranken, Beieren, heeft nog dezelfde Y-DNA clade. Bronstijd DNARond 6.800 jaar terug zien we bij het begin van de Bronstijd opnieuw een omslag. Haplogroep G wordt in korte tijd grotendeels verdrongen door een volk van zeer succesvolle metaalbewerkers en paardenfokkers. Zij hebben de Y-DNA haplogroep R. De noordelijke tak R1a, de zuidelijke tak R1b. Het zijn de dragers van de Touwbeker of strijdhamer cultuur van 5000 tot 4300 voor heden. Zij spreken proto-Germaans. De vraag is of hier sprake was van Genocide of Verdringing. Zeker is ook dat pest epidemieën zowel een oorzaak van de volksverhuizingen geweest kan zijn. Bij de residente bevolkingen leidde dit tot grote sterfte . Wel blijkt dat de helft van de huidige Europese bevolking bestaat uit afstammelingen van slechts enkele mannen, koningen met Y-DNA aan het begin van haplogroep R. IJzertijd DNAOngeveer 4.000 jaar geleden, aan het begin van de IJzertijd, doet zich weer een dramatische verandering voor in het Y en mt-DNA van Europa. Dit was waarschijnlijk gerelateerd aan het begin van de droge tijd in het Midden-Oosten, na de lange natte periode op het Arabische schiereiland, die van 8500 tot 4500 jaar geleden duurde. Romeinse TijdEen veelheid aan Y-DNA claden ontstaat. In Europa is er toenemende welvaart en daarbij een grote mobilititeit in alle delen van de bevolking. * De dramatische verandering in het Y-DNA in het NeolithicumIn tegenstelling tot de demografische reconstructies op basis van mt-DNA, zien we een sterke tweede bottleneck (flessenhals) in de Y-chromosoom lijnen in de laatste 10.000 jaar. De veronderstelling is dat dit knelpunt veroorzaakt is door culturele veranderingen waarbij een slechts kleine groep mannen deel nam aan de voortplanting. We denken dan aan opgetreden sociale stratificatie met veelwijverij. De eerste flessenhals in de Y-lijnen was na het vertrek uit Afrika gevolgd door de snelle kolonisering van Eurazië in het korte tijdsinterval van 47-52.000 jaar geleden. Deze tweede dramatische afname is gevolgd door een spectaculaire groei van nieuwe Y-DNA subgroepen tussen 8000 en 4000 jaar terug. Het vrouwelijk DNA bleef daarentegen tot 17 maal hoger in diversiteit. (34) 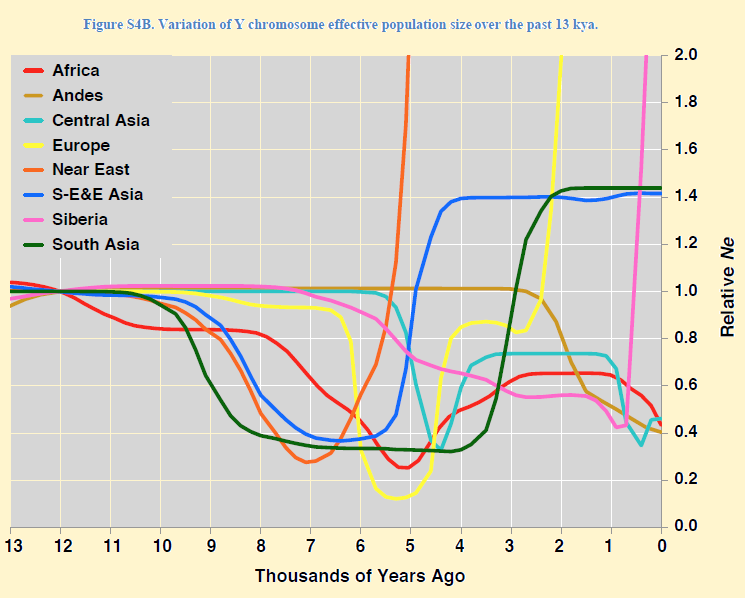
De dramatische gebeurtenissen voor ons mannen in het Neolithicum, de Brons- en IJzertijd. Age.
* Vervanging van het Mt-DNA in het Neolithicum
|
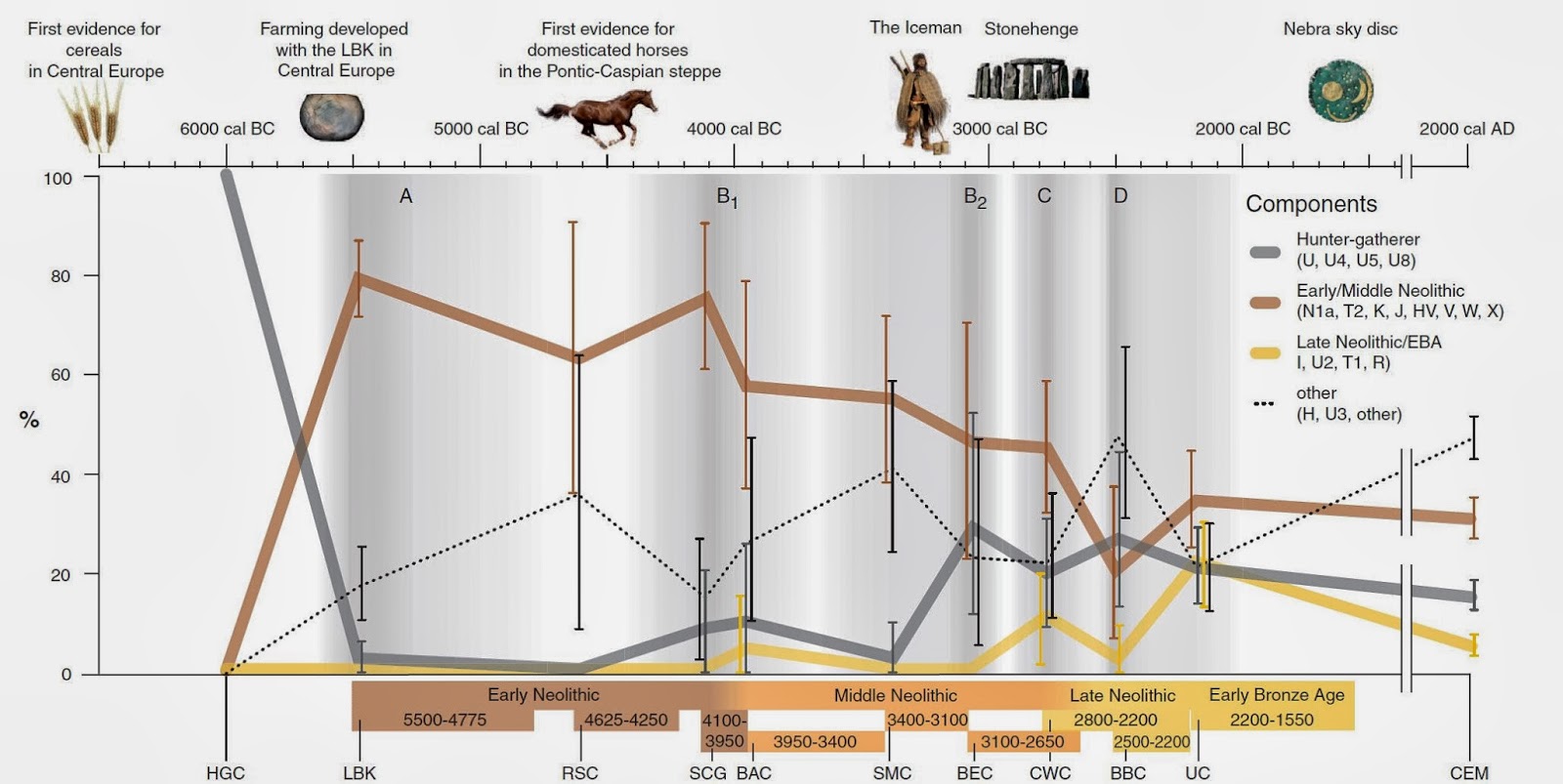
mt-DNA Tijdlijnen
* |
SamenvattingWij zien in de bevolkingsgeschiedenis van Europa de volgende elkaar opvolgende populaties: . |
|
Populatie 1. Neanderthalers.
|
Begintijd circa 200.000 jaar geleden
|
Eindtijd circa 44.000 jaar geleden
|
|
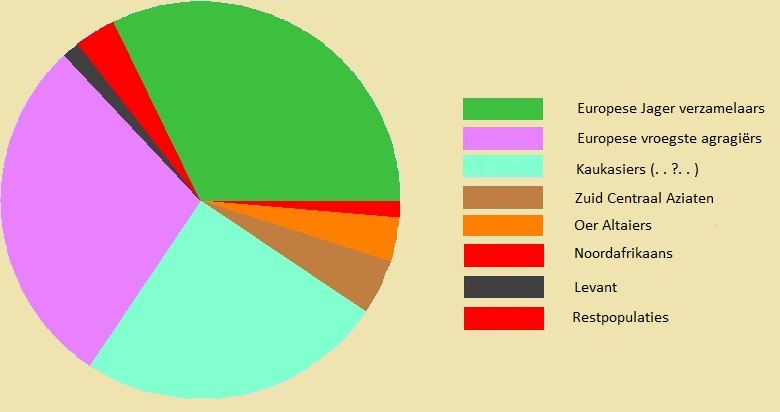
* Autosomaal DNA (AuDNA) in een Maastrichtenaar Bij deze Maastrichtenaar zijn de genen van alle volken van de Europese prehistorie aan te tonen.
|
|
Jager-verzamelaars
|
32%
|
 |
|
* Recente vervanging van het Y-DNA op CubaEen goed voorbeeld van de verdringing van de autochtone genen door een geheel nieuw genenpakket toont de genografische samenstelling van het Caribische eiland Cuba. Het eiland is al gedurende 7000 jaar bewoond. In 1513 werd het een Spaanse kolonie. Er woonden toen ongeveer 110.000 mensen. Nu zijn het er ruim elf millioen. De nieuwkomers waren Spanjaarden maar ook veel Afrikanen kwamen als slaven het land binnen. In 2012 is een admixture analyse gemaakt uit een groep van 1.019 aselectief gekozen inwoners. Hiervoor werd een mengsel van autosomale, Y-DNA en mt-DNA diagnostische haplogroep markers gebruikt. Zij vonden dat de oorspronkelijke Y-DNA haplogroepen bijna geheel verdwenen zijn. en wel voor 99½. Van de mt-DNA haplogroepen verdween twee derde. Bij de geïmporteerde bevolking van de zwarte slaven heeft dit proces zich ook, maar in veel geringere mate voorgedaan. Hun Y-DNA aandeel is de nu helft van het verwachtte percentage, dit in vergelijk met hun mt-DNA percentage van bijna 100%. (39) De conclusie die we hieruit kunnen trekken is dat bij een betrekkelijk vreedzame overheersing het mannelijk DNA nagenoeg geheel verdwijnt, maar het vrouwelijk DNA voor een groot deel behouden blijft. Wanneer dat laatste ook verdwijnt zal sprake zijn van uitroeiing. 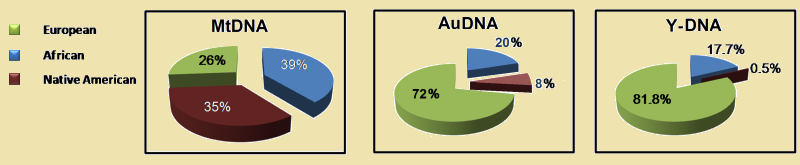
|
|
|
Mt-DNA
|
Autosomaal DNA
|
Y-DNA
|
|
Voorbeeld: Dit onderzoek heeft een betekenis die veel verder gaat dan dit eiland alleen. Dominante nieuwe bevolkingsgroepen zullen zich overal in de wereld en in alle tijden met vrouwen van onderworpen volkeren hebben voortgeplant. Van de onderworpen mannen zal zich slechts een klein deel hebben weten te handhaven. Oude kleine Y-DNA haplogroepen, zouden zo resten kunnen zijn van een oorspronkelijke bevolking. Haplogroep G is mogelijk een rest van zo'n oude onder de voet gelopen autochtone bevolkingsgroep. |
|
* Nederlandse DNA projectenZonen van AdamDit was een initiatief van het Nederlands Genootschap voor Geslacht en Wapenkunde (NGGW) en de Nederlandse Genealogische Vereniging (NGV). Opgezet in 2007 Vierhonderdzeventig mannen deden mee aan dit onderzoek, er werden 18 STR-markers onderzocht. De haplotypen en haplogroepen van de eerste groep deelnemers, ongeveer de helft, zijn gepubliceerd in Zonen van Adam in Nederland, Sahar Barjesteh van Waalwijk van Doorn-Khosrovani, Leo Barjesteh van Waalwijk van Doorn, Toon van Gestel en Frans Plooij, 2008. Van de tweede groep zijn de uitslagen nooit gepubliceerd. Van zowel de deelnemers van de eerste reeks testen van 2007 en tevens van degenen die zich later lieten testen, maar wier resultaten nog op publicatie wachten staan in deze volledige deelnemerslijst. Oud Hertogdom Brabant ProjectDit is een Belgisch project waar ook Nederlanders aan mee kunnen doen. Er worden standaard 37 markers getest. Een Spreadsheet met een lijst van personen die hun Y-DNA gemeten hebben via Oud Hertogdom Brabant en met ook degenen die vermeld staan in Zonen van Adam in Nederland, en waarbij tevens verwijzingen naar FTDNA websites staan vermeld, geeft een goede mogelijkheid om een Y-DNA verwant in de Benelux te vinden. Het is te vinden op: Lijst . De tweede groep van Zonen van Adam is hier nog niet vermeld. De niet gepubliceerde uitslagen bevinden zich bij Leo Barjesteh. Het Project Oud Hertogdom Brabant loopt verder onder de naam: DNA-Projecten - Familiekunde Vlaanderen vzw Brabant - België - Open Project - Benelux. FTDNADe meesten beginnen hun DNA onderzoek bij de grote Amerikaanse firma FTDNA. Mannen starten met een reeks Y-DNA STR-markers. Een snelle screening kan al gedaan worden met 12 STR-markers. Hiermee wordt ook de haplogroep ongeveer aangeduid. Een test met 37 markers geeft een duidelijk beeld. Voor een precieze plaatsing in de Y-DNA stamboom zijn SNP testen nodig. Hierbij wordt de haplogroep exact vastgesteld. Wanneer de hoofdgroep bekend is wordt met de groepscoördinator overlegd welke SNP's bepaald kunnen, of moeten, worden om de precieze subgroep te bepalen en daardoor de juiste plaats in de Y-DNA stamboom te krijgen. De resultaten van FTDNA zijn helder en overzichtelijk op afzonderlijke haplogroeplijsten gepubliceerd. Er zijn ook landelijke en ethnische lijsten. Voor ons zijn interessant buiten de haplogroeplijsten: |
*
Op zoek naar onze antropologische achtergrond met DNA projectenMet behulp van data in grote databases, kan men de landen ontdekken waarin een bepaalde haplogroep (of beter een subgroep) het meest voorkomt. Naarmate van meer mensen, en groepen van mensen, de DNA-kenmerken bekend worden, zal men een beter beeld krijgen van de wereldgeschiedenis. Nu dagen de contouren al op hoe sommige subgroepen, claden, gehecht kunnen worden aan bepaalde oude volkeren, zoals Kelten, Friezen, Franken, Bourgondiërs, Saksen, Vikingen, Vandalen, Alanen, Sarmaten Hunnen, Lombarden, etc. FTDNA ‑ the Big Y projectThe Big Y test van het Y chromosoom ruim 60% van de SNP's. Hij test rond de 15 millioen plaatsen. De eerste resultaten zijn nu bekend. Full Genomes Sequencing De Full Genomes Sequencing is een test van zogenaamde hoge dekking. Ruim 90 % van het Y-chromosoom wordt afgelezen. Dit is op zo'n 25 millioen plaatsen.
* |
|
Auteur: Boed Marres |
Laatste bewerking: 1 januari 2019. |